 |
ReoID Phylogenetic sequence analysis and improved diagnostic assay systems for viruses of the family Reoviridae
Edited by Peter. P. C. Mertens and Houssam Attoui |
 |
| ||||||
|
Summary of the molecular epidemiology of Bluetongue virus in Europe since 1998: introductions, origins and movements of individual virus strains: Peter Mertens: Updated 25th July 2007 (For additional information (see Outbreak data and www.iah.bbsrc.ac.uk/bluetongue/)
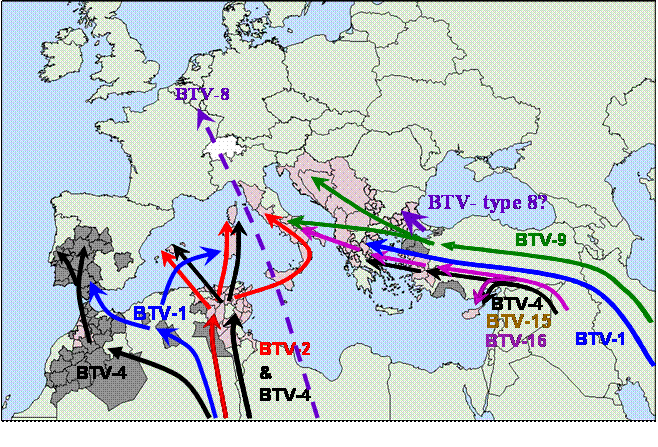
Figure 1: Genetic analysis of bluetongue viruses isolated in Europe has shown that six types of the virus (1, 2, 4, 8 9 and 16) have entered the region since 1998. There are four distinct routes by which these viruses have arrived. From the east via Turkey / Cyprus; from North Africa (Algeria, Tunisia) into Italy and the eastern Mediterranean Islands; from Morocco into southern Spain and Portugal. And via an unknown route into northern Europe. BTV-15 was identified in Israel during 2006, and represents a strain that may threaten Europe in the future . Further information is available from the dsRNA virus web site at : www.reoviridae.org/dsRNA_virus_proteins/
By combining data from disease surveillance and diagnostic testing, with the results of molecular epidemiology studies, it is possible to build up a picture of the recent colonisation of Europe by bluetongue virus and its subsequent spread in the region . Genome segment 2 encodes the virus outer coat protein VP2, which is the most variable virus protein and is the primary determinant of virus serotype. The majority of the data regarding the molecular epidemiology of European BTV strains were generated for genome segment 2, although data was also produced for segments 5, 6, 8 and 10.
BTV-1: www.reoviridae.org/dsRNA_virus_proteins/ReoID/btv-1.htm There have been two separate introductions of BTV-1 into the Mediterranean region which have occurred in 2001 and 2006. These are represented in the reference collection by the Greek isolate GRE2001/01 and by Algerian and Moroccan isolates ALG2006/01 and MOR2006/06 (report: Typing of BTV-1 from Morocco 2006). Phylogenetic analyses of segment 2 sequences not only identified these strains as BTV serotype 1 but also showed that they are distinct, belonging to different ‘eastern’ and ‘western’ lineages respectively (www.reoviridae.org/dsRNA_virus_proteins/BTV1-segment2-tree.htm). The initial strain of BTV-1 in Greece is related to eastern viruses from India and Malaysia and is thought to have entered Europe from the east, most probably from Turkey. This strain only persisted for a relatively short period in Greece and did not spread to other European Countries. In 2006 BTV was detected in northern Africa and was shown by RT-PCR and sequence analysis to be caused by BTV type 1. The strain involved is most closely related to other western type 1 isolates from sub-Saharan Africa, but is clearly distinct from the South African vaccine strain (RSAvvvv/01). It is also distinct from the earlier Greek isolates of BTV-1. There are subsequent reports of BTV-1 in Sardinia during 2006, suggesting that the virus had moved northwards into the western Mediterranean Islands. In July 2007 BTV-1 was identified in sheep with clinical signs of disease (which had previously been vaccinated against BTV-4) in the South of the Iberian peninsula (Spain).
BTV-2: www.reoviridae.org/dsRNA_virus_proteins/ReoID/btv-2.htm BT outbreaks were recorded in Tunisia during 2000. The virus was isolated and is represented in the IAH reference collection by TUN2000/01 and TUN2000/02. Phylogenetic analyses of Seg-2 sequences identified the virus as BTV serotype 2, belonging to a western lineage (www.reoviridae.org/dsRNA_virus_proteins/BTV2-segment-2-tree.htm). The virus is related to other BTV-2 strains from Nigeria and South Africa, although it is distinct from the South African BTV-2 vaccine strain (RSAvvvv/02). The initial Tunisian isolates of BTV-2 are also almost identical to subsequent isolates made in Corsica and Sardinia indicating that (like the BTV-1 strain from North Africa in 2006) BTV-2 managed to spread northwards, from north Africa into Italy and the western Mediterranean islands (represented by isolates from Corsica (FRA2001/01 to FRA2001/06), Sardinia (SAD2001/01 to SAD2002/03) and Sicily (ITL2002/01). However, there is evidence that some of the subsequent isolates of BTV-2 from mainland Italy, are similar to the BTV-2 vaccine strain that was also used in the region, indicating that it can be transmitted in the field.
BTV-4: www.reoviridae.org/dsRNA_virus_proteins/ReoID/btv-4.htm Widespread bluetongue outbreaks in Greece during 1999 and 2000 were caused by a strain of BTV-4 (represented in the collection by multiple isolates: GRE1999/01 to GRE2000/07). Phylogenetic analyses demonstrated that the virus is similar to earlier isolates from Cyprus and Turkey (CYP1969/01 and TUR1978/01) (www.reoviridae.org/dsRNA_virus_proteins/btv4-segment-2-tree.htm). It is also closely related to the Reference strain of BTV-4, cited in the collection as RSArrrr/04 , which is also believed to have been originally derived from Cyprus. This suggests that the BTV-4 strain which invaded Greece and the Eastern Mediterranean region since 1999, has been circulating in the region, on the borders of Europe, for some time. Indeed several isolates of BTV-4 have subsequently been made in Israel (represented by isolates ISR2002/01 to ISR2001/13, ISR2006/10 and ISR2006/12). Phylogenetic analyses show that these European BTV-4 strains belong to a western lineage but are distinct from the South African vaccine strain (represented in the collection by TURvvvv/04). In 2003, BTV was detected in the Western Mediterranean islands, including Menorca (represented in the reference collection by isolates SPA2003/01 to SPA2003/04. Although phylogenetic analyses showed that this was also a western strain of BTV-4, it is clearly distinct from strains that had previously caused outbreaks in the eastern Mediterranean region. These conclusions were confirmed by Stephan Zientara et al, working with Corsican isolates at Maisons Alfort in France. This virus is believed to have entered Europe from North Africa, possibly (like the earlier BTV-2 strain) from Tunisia. The same strain of BTV-4 subsequently caused outbreaks and was isolated in Morocco (represented by isolate MOR2004/02) and then spread to the Iberian peninsula in 2004 (represented by isolates SPA2004/01 and SPA2004/02), where it persisted through into 2005 (represented by isolates SPA2005/02 to SPA2005/05) Three virus-isolates were made in Israel from outbreaks of disease in 2006. One of these was typed as BTV-15 (see below) while the other two were typed as BTV-4, in RT-PCR assays using serotype specific primers targeting genome segment 2 (represented by strains ISR2006/12 and ISR2006/10). (report: Typing of BTV4 and BTV-15 isolates from Israel 2006).
BTV-8: www.reoviridae.org/dsRNA_virus_proteins/ReoID/btv-8.htm In August 2006, BT was recognised in the Netherlands (the first time in northern Europe) as classical clinical signs in infected sheep. This outbreak was not anticipated, and its rapid recognition is a reflection of the vigilance and effectiveness of local veterinary services in the Netherlands . The disease was subsequently detected in Belgium (where the initial infection is thought to have started - the index case, possibly in May or June 2006), Luxemburg, Germany and north-east France. Multiple isolates of the virus were made from across this region (represented in the collection by NET2006/01 to NET2006/04, GER2006/01 and BEL2006/01). Sequence analyses of Seg-2 from NET2006/01, demonstrated that the virus is from a western lineage from sub Saharan Africa, but is distinct from the BTV-8 vaccine strain (www.reoviridae.org/dsRNA_virus_proteins/BTV-8-Seg-2-tree.htm) This was confirmed by full genome sequence analysis of NET2006/04 [4] (see typing reports: Identification of BTV-8 in the Netherlands 2006 also Comparison of genome segment 2 sequences of BTV-8 isolates). It is uncertain exactly how BTV-8 arrived in northern Europe, but the absence of BTV-8 outbreaks in southern Europe suggests that it did not involve simple linear extension of earlier outbreaks and is likely to reflect a distinct entry route and mechanism. Interestingly serological evidence for BTV-8 infection of sentinel animals was subsequently obtained from Bulgaria (during 2006), although since no virus was detected by virus isolation or by RT-PCR assays of blood samples, it is difficult to determine the likely timing of BTV infections involved or the relevance of these reports to northern Europe (report Identification of antibodies to BTV-8 (but not virus) in Bulgaria 2006). In April 2007 a sentinel animal in Germany seroconverted, becoming seropositive for BTV. This suggests that the virus had not 'died out' during the winter. The virus was identified in Germany by type specific RT-PCR, as BTV-8 (isolates GER2007/01 to GER2007/04). On July 17th 2007 the Belgian authorities also reported disease in a flock of sheep near Antwerp in which a single sheep had clinical signs of bluetongue and subsequently died. The detection of rising levels of antibodies to BTV and viral RNA in blood samples indicated the presence of virus and new transmission in 2007. New cases have also been reported during July in the Netherlands and in north east France. During August 2007 there was a massive increase in the number of outbreaks across the entire region, including notably an increase in Northern France, which experienced only a small number of cases in 2006.
BTV-9: www.reoviridae.org/dsRNA_virus_proteins/ReoID/btv-9.htm The first serotype to arrive in the Europe in 1998 was BTV-9 (represented by virus isolate GRE1998/01). Sequence data for genome segment 2 of this isolate was compared to other isolates of BTV held in the reference collection at IAH Pirbright from around the world (see www.reoviridae.org/dsRNA_virus_proteins/ReoID/btv-9.htm). The resulting analysis shows that this is an ‘eastern’ virus, related to strains from Indonesia and Australia (www.reoviridae.org/dsRNA_virus_proteins/btv9-segment-2-tree.htm). This strain of BTV is also closely related to BTV-9 subsequently isolated in Greece, Bulgaria, Turkey, Bosnia, Kosovo and Serbia, represented in the reference collection (www.reoviridae.org/dsRNA_virus_proteins/ReoID/btv-9.htm) by multiple isolates from these regions. This indicates that the outbreak of BTV-9 in the eastern and central Mediterranean region stemmed from this eastern source. It also appears likely that BTV-9 in Italy originated from the same initial source (since no other introductions of BTV-9 field strains have been detected), although an isolate of BTV-9 was made in Sicily (ITL2003/01) which was identical to the BTV-9 vaccine strain.
BTV15 www.reoviridae.org/dsRNA_virus_proteins/ReoID/btv-15.htm A sample of a virus-isolate that was made in Israel from outbreaks of disease in 2006 (represented in the collection by ISR2006/11) was typed as BTV-15, by RT-PCR assays using serotype specific primers targeting genome segment 2 . Subsequent phylogenetic analyses confirm the virus serotype and indicated that it belongs to a western lineage (www.reoviridae.org/dsRNA_virus_proteins/BTV15-Seg2-tree.htm). The number of BTV-15 isolates that are available is very limited. So it is difficult to be more precise about the origins of this virus strain. However, the existing data suggest that it is new to the region and may therefore represent a further threat to Europe in the future (report: Typing of BTV4 and BTV-15 isolates from Israel 2006).
BTV16: www.reoviridae.org/dsRNA_virus_proteins/ReoID/btv-16.htm The initial European strain of BTV-16 was isolated in Greece during 1999 (represented in the reference collection by isolate number GRE1999/13). Sequence analysis of Seg-2 and phylogenetic analysis (www.reoviridae.org/dsRNA_virus_proteins/btv16-segment2-tree.htm) show that this virus is from an 'eastern' lineage but is very similar to strains of BTV-16 from Turkey and the South African reference strain of BTV-16. This may seem surprising but the reference strain (RSArrrr/16) was originally derived from an outbreak in Pakistan. This suggests that the virus arrived in Europe from the east, possibly via Turkey. Sequencing studies have shown that both the Turkish and Greek strains are also very losely related to the strain from Cyprus (CYP2004/01), suggesting that these eastern Mediterranean strains represent a single common lineage. Phylogenetic analyses have shown that the original European field strain of BTV-16 is closely related, although distinct from the BTV-16 vaccine strain. Its appearance in Europe may be related to the use of live BTV-16 for several years as part of an annual vaccination campaign in Israel. After its arrival in Europe the virus spread eastwards across Europe and although BTV-16 eventually appeared in mainland Italy in 2002, this strain appears to be more closely related to the vaccine strain (~99.9% in Seg-2) suggesting that it was not derived from the earlier Greek or Turkish outbreaks, although at that stage the BTV-16 vaccine had not yet been used in Italy. BTV-16 also caused an outbreak in Sardinia in 2004, and the strain involved (represented by isolates SAD2004/01 to SAD2004/24) is identical to the BTV-16 vaccine that was used in mainland Italy, suggesting that this was a ‘vaccine’ outbreak and again was not caused by the European field strain represented by viruses from Greece, Turkey and Cyprus.
Summary With the development of a molecular epidemiology database containing sequence data for large numbers of specific well documented BTV isolates, sequence analyses and comparisons of genome segment 2 can now be used to give a clear indication of the serotype and origins of individual BTV strains (Maan et al 2007). These analyses for the European BTV strains isolated since 1998 have indicated that six serotypes of the virus have entered Europe, with at least 9 separate introductions, via at least 4 distinct routes. Indeed there have been new introductions in each year with the possible exception of 2002. The use of live vaccine strains of BTV-2, 4, 9 and 16 in the region have added further strains to the European virus pool. With this number of distinct viruses co-circulating, it is perhaps not surprising that there is also conclusive evidence for genome segment exchange via ‘reassortment’ events between different virus strains and serotypes, and between vaccine and field strains, in the field (Batten et al manuscript in preparation). There appears to have been a fundamental change in the European ecosystem since 1998, possibly linked to climate change (Purse et al 2005) which favours the introduction and survival of BTV in Europe. Based on these recent events, it considered likely that other BTV strains, and possibly other orbiviruses, will spread to the region in the immediate and foreseeable future.
References 1. Bethan V. Purse, Philip S. Mellor, David J. Rogers, Alan R. Samuel, Peter P. C. Mertens & Matthew Baylis (2005) Climate Change And The Recent Emergence Of Bluetongue In Europe Nature Reviews Microbiology 3, 171-181. 2. Maan S., Maan N.S, Samuel A.R., Rao S, Attoui, H., & Mertens P.P.C (2007) Analysis and Phylogenetic Comparisons of Full-Length VP2 Genes of the Twenty-Four Bluetongue Virus Serotypes. Journal of General Virology (in Press). 3. Breard E, Sailleau C, Nomikou K, Hamblin C, Mertens PP, Mellor PS, El Harrak M, Zientara S. (2007 ) Molecular epidemiology of bluetongue virus serotype 4 isolated in the Mediterranean Basin between 1979 and 2004.Virus Res.125, 191-197. Mertens, P.P.C. Maan N. S., Prasad, G., Samuel A.R., Shaw A., Potgieter, A.C., Anthony, S. J., and Maan S. (2007) The design of primers and use of RT-PCR assays for typing European BTV isolates: Differentiation of field and vaccine strains Journal of general Virology (in Press) 4. S. Maan, N.S. Maan, N. Ross-Smith, C.A. Batten, A.E. Shaw, S.J. Anthony, A.R. Samuel, K.E. Darpel, E.Veronesi, C.A.L. Oura, K.P. Singh, K. Nomikou, A.C. Potgieter, H. Attoui, E.van Rooij, P. van Rijn, K. De Clercq, F.Vandenbussche, S. Zientara, E.Bréard
C. Sailleau, M.Beer, B. Hoffman, P.S. Mellor, P.P.C. Mertens (2008) Sequence analysis of bluetongue virus serotype 8 from the Netherlands
2006 and comparison to other European strains. Virology 377, 308-318.
|
|
|